Skjult informasjon på linje med et isfjell?
(Fra: The Mysterious Epigenome; Hva ligger bortenfor DNA; Th.E.Woodward; J.P.Gills; Kregel Publications; 2012)
For å ta tak i det sentrale spørsmål om opprinnelsen til ny informasjon i DNA, trenger vi å gjenfortelle historien om det menneskelige genom (‘human genome project’) i USA. På sommeren i år 2000 ble det holdt en pressekonferanse, for å annonsere at en milepel var nådd i genetikkens historie. Etter at ett nettverk av forskere gjennom et tiår hadde utforsket genenes plassering i det menneskelige genom, ble det hold en pressekonferanse, initiert av presidenten. De hadde funnet den presise ‘bokstav-for-bokstav’ sammensetning for de i alt 3,2 milliarder bokstaver i det menneskelige genom. Daværende leder for prosjektet, Fr. Collins, hadde utarbeidet talen holdt av president Clinton. Der skrev han: «I dag lærer vi språket, hvorved Gud skapte livet.. Det er ydmykende og fyller meg med ærefrykt at vi har fått det første glimtet av vår egen instruksjonsbok, tidligere bare kjent av Gud.»
 Bilde 1 Odds mot menneskelig evolusjon
Bilde 1 Odds mot menneskelig evolusjon
Fr.Collins hadde en fortid som ateist, og blitt en evangelisk kristen i 1993. Senere skrev Collins en bok med navn, ‘The Language of God’ (2006). Så langt Fr. Collins. Det manglet mye på at det var gjengs syn blant biologene på prosjektet, som mente talen og boka bare var uttrykk for Collins’ eget syn. Det hadde i og for seg vært greit, om de bare vedkjente det samme, for sitt motsatte syn. At prosjektet hadde kostet 3 milliarder dollar av skattebetalernes penger, var også et poeng i USA.
‘Slutning til Design’
(Intelligent Design –ID)
Det er mer enn poetisk likhet mellom ‘Guds språk’ og DNA-koder, som kodes til RNA og oversettes til proteinkjeder. Det er fullt mulig å se at kodene, språket og pakkene med informasjon i cellene, sender oss et ‘signal om intelligens’. Det innebærer at de presenterer et unikt mønster for oss, som ellers i universet; eks. NASA’s ExtraTerrestrial Intelligence (ETI) program, der kodede signaler fra verdensrommet evt. henføres til et intelligent opphav. ID-bøker som ‘Signature in the Cell’ (S.C.Meyer;2009) og ‘The Edge of Evolution’ (M.Behe; 2007) gir eksempler på dette. Men noen motstandere av ID har sett på ID som ‘kreasjonisme i billig smoking’, og betraktet det som en fare for USA’s ledende rolle innen vitenskap, ja som ‘en fare for vitenskapens framtid og vestlig sivilisasjon’..
Det er litt historieløst, i og med at ID benytter et prinsipp fra Darwins nære venn og med-vitenskapsmann, den britiske geologen Ch. Lyell. Det er prinsippet om uniformitarisme, som går på den (omstridte) holdning at nåtiden er nøkkelen til fortiden. En trenger å være kritisk i bruk av regelen, som har sine begrensninger, men den har likevel blitt brukt som en vitenskapelig nøkkel til fortiden. Her er hvordan ID benytter prinsippet: ‘Om en i nåtiden ser at kodede signaler og språklige informasjons-pakker, stammer fra intelligente årsaker, så er det grunn til å henføre samme type virkning (DNA’s digitale database) til samme type årsak -Intelligens, også i fortiden.
Om vi ser enkle meldinger, formet i SCRABBLE-bokstaver, f.eks: «Er og svømmer, kan du lufte hunden?» - en enkel melding på 36 tegn; så skal det litt uvilje til å tolke det som at klossene ramlet tilfeldig ned slik. Den enkleste, kjente bakterie (Nanoarchaeum equitans) har et genom på 490.885 DNA bokstaver, presis organisert i 552 gener.»
En genetisk oppvekker
Det som gir grunnlag for ‘slutning til Design’ når det gjelder 36 bokstaver hjemme på kjøkkenbordet, eller ca. en halv million DNA-bokstaver i enkleste bakterie-genom, burde gjelde desto sterkere, om hele DNA-biblioteket i planter og dyr ble funnet å være tett integrert med et høyere ordens informasjons-og-kontroll system. DNA synes nå å være cellens ‘arkiv’, billedlig talt, med ‘skuffer som inneholder kategoriserte dokumenter’. Avhengig av celletypen, vil visse skuffer åpnes ofte, og kopieres og tas over til mangfoldiggjøring. Hele prosessen ‘å avgjøre hvilke skuffer som skal åpnes og kopieres, til rett tid styres av epigenetikken. Den koordineres av multiple algoritmer, for å framskaffe enormt kompliserte ‘byggverk’ og kontrolloperasjoner. Det er en overordnet prosess som en nå har begynt å få dypere innsyn i.
Bilde 2 Celle-detaljer
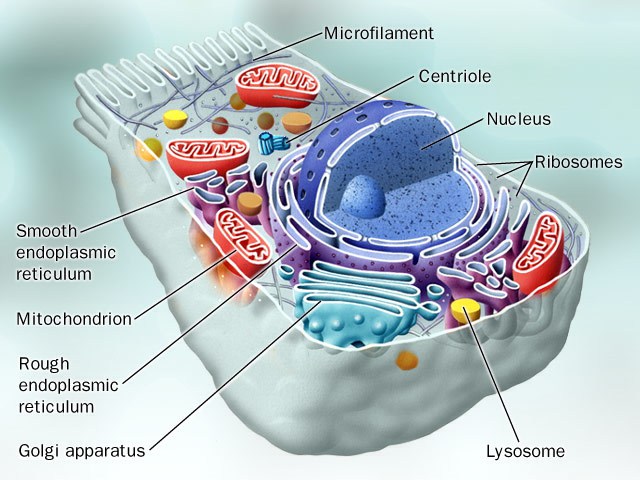
Det er her det nå føles en del vitenskapelige ‘rykninger’. Vi fortalte i forrige avsnitt om biologenes store overraskelse, i det de avdekket lag på lag av styrende tilleggsinformasjon, utover det genetiske arkiv-systemet med DNA. Denne tilleggsinformasjonen kalles gjerne epigenetikk, etter epi: over/overfor genomet. Epigenomet henfører til hele det koordinerte kontroll-systemet som befinner seg utenom DNA-sekvensen. Forskning fokuserer i stigende grad på disse styrende informasjonslag {-som billedlig kan sammenlignes med et SQL-styrt databaseverktøy som virker på det filbaserte DNA-arkivet-oversetters bildebruk}.
R. Sternberg, som innehar doktorgrader i evolusjonsbiologi og system-biologi, har forsket mye på såkalt ‘junk-DNA hypotesen’, og kritisert denne. Han har bekreftet at utenom og ovenfor DNA har forskere snublet over ‘lag på lag’ med ny cellulær informasjon. I samtale med ham, ble det foreslått en metafor: at det vi kan se av DNA i cellekjernen, svarer til hva en kan se av et isfjell i havet. I en artikkel i Time, ble genomet avtegnet som en prikk, mens epigenomet, relativt -i forhold til genomet, var som en sirkel med diameter på mer enn tre tommer. Hvor vi finner epi-genomet, hvordan det virker og hva dets rolle er, skal vi se nærmere på i det vi skal dypere inn i menneske-cellen.
Epigenetisk tagging
DNA’s 4 bokstaver + 1
(Nøkkel til kreftens gåte?)
 Bilde 3 Metylering (blåfarget) av Cytosin i DNA
Bilde 3 Metylering (blåfarget) av Cytosin i DNA
En kan tenke på en metyl-tag som ett nedstrippet metan-molekyl (ett H-atom mindre). I dyr er det C(ytosin) som tagges med metyl. Celler vil noen ganger ha metyl-tagging på noen av sine C(ytosin) bokstaver. Metyl-tagging er formet som små hoder med 3 øreformede utvekster. Metylering spiller en viktig rolle i epigenomets regulering av vekst og ved sykdom. Hypermetylering kan lede til kreft. I senere år har en oppdaget at dette mønsteret av tagging er vitalt for cellens funksjonsevne. Metylering av utvalgte C-bokstaver kan være den mest massive og mest mystiske, i alle nivåer til epigenomet. Titalls-millioner av trinn i DNA-et er tagget på dette viset. Noen regner metylert-C som DNA’s 5.bokstav, i tillegg til A, T, G og C.
Andre markører i epigenomet
Vi har diverse markører i epigenomet:
i)
Det er funnet at festestedet for mikrotubuler og celleskjelett i cellemembranen er plassering for en bestemt type av epigenetisk informasjon.
ii) Histoner kodes med med en egen acetyl-kode, på et helt spesifikt sted på histonet.
Bilde 4 Histonspoler merket med acetyl
iii) Vi har også 3 andre former for tagging: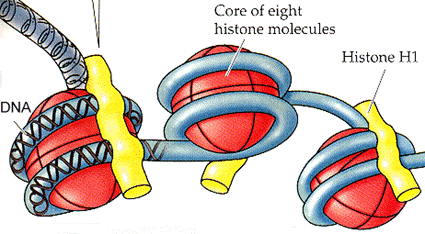
a. Fosfat-tagger. Disse små molekylene er litt større enn metyl-tagger. De benyttes til å bygge sidetrinnene til DNA og RNA.
b. Ubiquitin proteiner. Disse er velkjente og betydelig større enn før nevnte tagger. De knyttes gjerne til ødelagte eller utslitte deler av cellen, for å merke dem for ødeleggelse.
c. SUMO (Small Ubiquitin-like MOdifiers)-proteiner er på størrelse med ubiquitiner. Dette er protein maskiner med nærmest ‘gåseliknende’ form, benyttes til å merke diverse proteiner for ulike formål.
Det finnes mange flere forhold ved epigenomet som vi kommer tilbake til. Særlig viktig er et mystisk, men viktig system som kalles zygote-koden.
Rollen til epigenom-systemet
Dette er under stadig utforsking, men data synes å tyde på at nærvær av tagger ( enten inni genet, nær endene eller genets tilgrensende område) synes å ‘stilne’ dette genet. Om det ikke er metyl-tagger knyttet til det, er det ‘skarp ammunisjon’ klar til å benyttes på ett øyeblikks varsel. Et metyl-tagget gen er som en pasient, gitt et bedøvende middel, for å stille ham til ro. Disposisjon av DNA kontrolleres ved kjemiske signaler innen tagging av spolen. Acetyl-tagging synes å åpne histon-spolene, mens metyltagging synes å lukke til/sikre spolene. Fosfat-tagging på histon-spolenes hale og linking til SUMO-proteiner (eller ubiquitin proteiner) på siden av spolene, synes også å påvirke hvorvidt DNA er åpen eller låst for bruk.
Bilde 5 Gåten metylerte gener
(en av kreftens skjulte årsaker)?
Om ett spesielt gen hjelper å regulere celle-deling, så liter cellen på at det genet forhindrer at celle-deling kommer ut av kontroll. Om halen på histon-proteiner ‘klemmer’ DNA veldig stramt, sier Dr. Issa at genet er skjult for cellen. Et gen som er skjult kan ikke bli benyttet. Det har samme effekt som om genet er dødt eller mutert. Dette er forhold som regulerer gen-uttrykk og kan løpe løpsk ved ikke-styrt celledeling (kreft). I dag kjenner man til at redusert eller økt metylering, kan virke som triggere for visse kreftformer. F.eks. kan oncogener, som normalt er ‘velvillige’, spille en nøkkel rolle i at en normal celle forvandles til en kreftcelle, når de muteres eller overeksponeres. I noen tilfelle har en funnet at normal ‘stilning’ av slike oncogener, blir forstyrret av feilaktig C-tagging av DNA-bokstavene. Når cellen er programmert til å fase ut i normal status, gjennom normal celle-død (apoptosis), kan fjerning av metyltagging gjøre at oncogener slås på og ‘spinner vilt’. I andre tilfelle kan det være uønskede metyltagger, som legges til kreft-undertrykkende gener og hindrer dem fra å gjøre tiltenkt jobb.
Den sentrale jobben til epigenomet er å kontrollere hvordan DNA uttrykkes, med sine tusener av gener i ulike celletyper. Hvordan den får utøve denne rollen har sammenheng med kreft og andre sykdommer, som vi kommer tilbake til.
Oversatt og tilpasset .htm-format ved Asbjørn E. Lund
{kind=link}